



一、Підвищити чутливість реакційної системи:
1. Виділення високоякісної РНК:
Успішний синтез кДНК відбувається з високоякісної РНК.Високоякісна РНК повинна бути принаймні повнорозмірною та не містити інгібіторів зворотної транскриптази, таких як EDTA або SDS.Якість РНК визначає максимальну кількість інформації про послідовність, яку ви можете транскрибувати в кДНК.Поширеним методом очищення РНК є одноетапний метод із використанням гуанідинізотіоціанату/кислого фенолу.Щоб запобігти забрудненню слідовими кількостями РНКази, РНК, виділену з багатих РНКазами зразків (таких як підшлункова залоза), необхідно зберігати у формальдегіді, щоб зберегти високоякісну РНК, особливо для тривалого зберігання.РНК, виділена з печінки щурів, в основному розкладалася після зберігання у воді протягом одного тижня, тоді як РНК, витягнута з селезінки щурів, залишалася стабільною після зберігання у воді протягом 3 років.Крім того, транскрипти, довші за 4 кб, більш чутливі до деградації слідовими РНКазами, ніж малі транскрипти.Щоб підвищити стабільність збережених зразків РНК, РНК можна розчинити в деіонізованому формаміді та зберігати при -70°C.Формамід, який використовується для збереження РНК, не повинен містити дебрисів, що розкладають РНК.РНК підшлункової залози може зберігатися у формаміді щонайменше один рік.Готуючись до використання РНК, ви можете використовувати такий метод для осадження РНК: додайте NaCl до 0,2 М і 4-кратний об’єм етанолу, помістіть при кімнатній температурі на 3-5 хвилин і центрифугуйте при 10 000×g протягом 5 хвилин.
2. Використовуйте RNaseH-неактивну (RNaseH-) зворотну транскриптазу:
Інгібітори РНКази часто додають до реакцій зворотної транскрипції для збільшення довжини та виходу синтезу кДНК.Інгібітори РНКази слід додавати під час реакції синтезу першого ланцюга в присутності буфера та відновника (такого як DTT), оскільки процес, що передує синтезу кДНК, денатурує інгібітор, таким чином вивільняючи зв’язану РНКазу, яка може розщепити РНК.Протеїнові інгібітори РНКази лише запобігають деградації РНК РНКазами A, B, C і не запобігають потраплянню РНКази на шкіру, тому будьте обережні, щоб не ввести РНКазу з пальців, незважаючи на використання цих інгібіторів.
Зворотна транскриптаза каталізує перетворення РНК в кДНК.І M-MLV, і AMV мають ендогенну активність РНКази H на додаток до власної полімеразної активності.Активність РНКази H і активність полімерази конкурують одна з одною за гібридний ланцюг, утворений між матрицею РНК і праймером ДНК або ланцюгом подовження кДНК, і розкладають ланцюг РНК у комплексі РНК:ДНК.Матрица РНК, деградована активністю РНКази, більше не може служити ефективним субстратом для синтезу кДНК, що зменшує вихід і тривалість синтезу кДНК.Таким чином, було б корисно усунути або значно зменшити активність РНКази H зворотної транскриптази.。
Зворотна транскриптаза SuperScript Ⅱ, зворотна транскриптаза RNaseH-MMLV і зворотна транскриптаза thermoScript, RNaseH-AMV, можуть отримати більшу кількість і більше кДНК повної довжини, ніж MMLV і AMV.На чутливість RT-PCR впливатиме кількість синтезу кДНК.ThermoScript набагато чутливіший, ніж AMV.Розмір продуктів RT-PCR обмежений здатністю зворотної транскриптази синтезувати кДНК, особливо при клонуванні більших кДНК.Порівняно з MMLV, SuperScripⅡ значно збільшив вихід довгих продуктів RT-PCR.РНКазаН-зворотна транскриптаза також має підвищену термостабільність, тому реакцію можна проводити при температурах, вищих за нормальні 37-42°С.За запропонованих умов синтезу використовуйте праймер оліго(dT) і 10 мкКі [α-P]dCTP.Загальний вихід першого пасма розраховували за допомогою методу осадження TCA.Повнорозмірну кДНК аналізували за допомогою сортованих за розміром смуг, вирізаних і підрахованих на лужному агарозному гелі.
3. Підвищте температуру інкубації для зворотної транскрипції:
Більш висока температура інкубації сприяє відкриттю вторинної структури РНК, збільшуючи вихід реакції.Для більшості матриць РНК інкубація РНК і праймерів при 65°C без буфера або солі з подальшим швидким охолодженням на льоду усуне більшість вторинних структур і дозволить праймерам зв’язатися.Однак деякі шаблони все ще мають вторинні структури, навіть після теплової денатурації.Ампліфікацію цих складних шаблонів можна здійснити за допомогою зворотної транскриптази ThermoScript і поставити реакцію зворотної транскрипції при вищій температурі для покращення ампліфікації.Вищі температури інкубації також можуть збільшити специфічність, особливо коли для синтезу кДНК використовуються ген-специфічні праймери (GSP) (див. Розділ 3).У разі використання GSP переконайтеся, що Tm праймерів збігається з очікуваною температурою інкубації.Не використовуйте оліго(dT) і випадкові праймери вище 60°C.Вибіркові праймери вимагають інкубації при 25°C протягом 10 хвилин перед підвищенням до 60°C.На додаток до використання вищої температури зворотної транскрипції, специфічність також можна покращити шляхом прямого перенесення суміші РНК/праймера з температури денатурації 65°C до температури інкубації зворотної транскрипції та додавання попередньо нагрітої 2× реакційної суміші (синтез кДНК з гарячим стартом).Цей підхід допомагає запобігти міжмолекулярному спаровуванню основ, яке відбувається за нижчих температур.Багаторазове перемикання температури, необхідне для RT-PCR, можна спростити за допомогою термоциклера.
Термостабільна полімераза Tth діє як ДНК-полімераза в присутності Mg2+ і як РНК-полімераза в присутності Mn2+.Його можна зберігати в теплі при максимальній температурі 65°C.Однак присутність Mn2+ під час ПЛР знижує точність, що робить Tth-полімеразу менш придатною для високоточної ампліфікації, такої як клонування кДНК.Крім того, Tth має низьку ефективність зворотної транскрипції, що знижує чутливість, і, оскільки зворотну транскрипцію та ПЛР можна виконувати за допомогою одного ферменту, контрольні реакції без зворотної транскрипції не можна використовувати для порівняння продуктів ампліфікації кДНК із забрудненою геномною ДНК.Продукти ампліфікації розділяли.
4. Добавки, що сприяють зворотній транскрипції:
До реакції синтезу першого ланцюга додають добавки, включаючи гліцерин і ДМСО, які можуть знизити стабільність подвійного ланцюга нуклеїнової кислоти та роз’єднати вторинну структуру РНК.Можна додати до 20% гліцерину або 10% ДМСО, не впливаючи на активність SuperScript II або MMLV.AMV також може переносити до 20% гліцерину без втрати активності.Щоб максимізувати чутливість RT-PCR у реакції зворотної транскрипції SuperScriptⅡ, можна додати 10% гліцерину та інкубувати при 45°C.Якщо до ПЛР додати 1/10 продукту реакції зворотної транскрипції, то концентрація гліцерину в реакції ампліфікації становить 0,4%, чого недостатньо для інгібування ПЛР.
5. Лікування РНКази H:
Обробка реакцій синтезу кДНК РНКазою H перед ПЛР може підвищити чутливість.Вважається, що для деяких матриць РНК у реакції синтезу кДНК перешкоджає зв’язуванню продуктів ампліфікації, і в цьому випадку обробка РНКазою H може підвищити чутливість.Як правило, обробка РНКази H необхідна при ампліфікації довгих повнорозмірних цільових матриць кДНК, таких як малокопійний туберозний шероз II.Для цього складного шаблону обробка РНКази H посилила сигнал, створений SuperScript II або кДНК, синтезованою AMV.Для більшості реакцій RT-PCR обробка РНКази H необов’язкова, оскільки стадія денатурації ПЛР при 95°C зазвичай гідролізує РНК у комплексі РНК:ДНК.
6. Удосконалення методу виявлення малих РНК:
RT-PCR особливо складна, коли доступна лише невелика кількість РНК.Глікоген, доданий як носій під час виділення РНК, допомагає збільшити вихід малих зразків.Глікоген, вільний від РНКази, можна додавати одночасно з додаванням Тризолу.Глікоген розчинний у воді і може зберігатися у водній фазі разом з РНК, щоб сприяти подальшому осадженню.Для зразків менше 50 мг тканини або 106 культивованих клітин рекомендована концентрація глікогену, вільного від РНКази, становить 250 мкг/мл.
Додавання ацетильованого BSA до реакції зворотної транскрипції за допомогою SuperScript II може збільшити чутливість, а для невеликих кількостей РНК зменшення кількості SuperScript II і додавання 40 одиниць інгібітора нуклеази RNaseOut може підвищити рівень виявлення.Якщо глікоген використовується в процесі виділення РНК, все одно рекомендується додавати BSA або інгібітор РНКази при використанні SuperScript II для реакції зворотної транскрипції.
二、Підвищити специфічність RT-PCR
1. Асинтез CND:
Синтез кДНК першого ланцюга може бути ініційований трьома різними методами, відносна специфічність яких впливає на кількість і тип синтезованої кДНК.
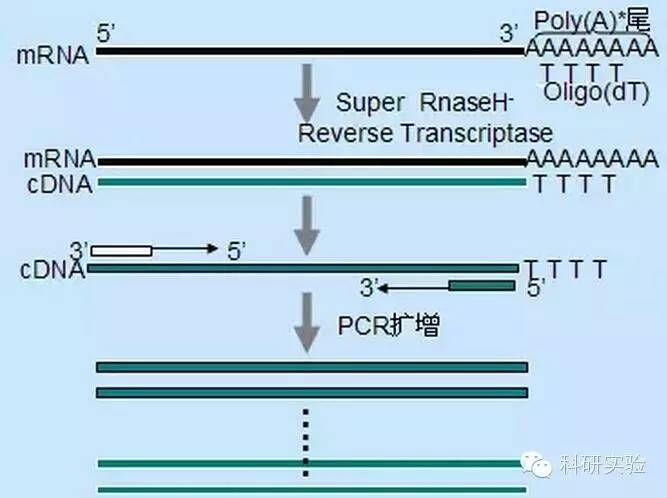
Метод випадкового праймера був найменш специфічним з трьох методів.Праймери відпалюються в кількох місцях транскрипту, утворюючи короткі кДНК часткової довжини.Цей метод часто використовується для отримання 5'-кінцевих послідовностей і для отримання кДНК з матриць РНК з областями вторинної структури або з сайтами термінації, які не можуть бути репліковані за допомогою зворотної транскриптази.Щоб отримати найдовшу кДНК, співвідношення праймерів і РНК у кожному зразку РНК потрібно визначити емпірично.Початкова концентрація випадкових праймерів коливалася від 50 до 250 нг на 20 мкл реакції.Оскільки кДНК, синтезована з сумарної РНК за допомогою випадкових праймерів, є переважно рибосомальною РНК, полі(А)+РНК зазвичай вибирають як матрицю.
Oligo(dT) праймери більш специфічні, ніж випадкові праймери.Він гібридизується з полі(А) хвостом, який знаходиться на 3'-кінці більшості еукаріотичних мРНК.Оскільки полі(А)+ РНК становить приблизно від 1% до 2% загальної РНК, кількість і складність кДНК набагато менші, ніж у випадкових праймерів.Через високу специфічність оліго(dT) зазвичай не потребує оптимізації співвідношення РНК до праймерів і відбору полі(А)+.Рекомендується використовувати 0,5 мкг оліго(dT) на 20 мкл реакційної системи.oligo(dT)12-18 підходить для більшості RT-PCR.Система ThermoScript RT-PCR пропонує оліго(dT)20 через його кращу термічну стабільність для вищих температур інкубації.
Геноспецифічні праймери (GSP) є найбільш специфічними праймерами для етапу зворотної транскрипції.GSP є антисмисловим олігонуклеотидом, який може специфічно гібридизуватися з цільовою послідовністю РНК, на відміну від випадкових праймерів або оліго(dT), які з’єднуються з усіма РНК.Ті самі правила, що використовуються для розробки праймерів ПЛР, застосовуються до розробки GSP у реакціях зворотної транскрипції.GSP може бути такою ж послідовністю, що й праймер ампліфікації, який приєднується до самого 3'-кінця мРНК, або GSP може бути сконструйований для відпалу після праймера зворотної ампліфікації.Для деяких ампліфікованих суб’єктів для успішної RT-PCR необхідно розробити більше ніж один антисмисловий праймер, оскільки вторинна структура цільової РНК може перешкоджати зв’язуванню праймера.Рекомендується використовувати 1 пмоль антисмислового GSP у 20 мкл реакції синтезу першого ланцюга.
2. Підвищте температуру інкубації для зворотної транскрипції:
Для повного використання всіх переваг специфічності GSP слід використовувати зворотну транскриптазу з вищою термостабільністю.Термостабільні зворотні транскриптази можна інкубувати при більш високих температурах, щоб підвищити жорсткість реакції.Наприклад, якщо GSP відпалюється при 55°C, специфічність GSP не буде повністю використана, якщо AMV або M-MLV використовується для зворотної транскрипції при низькій жорсткості 37°C.Однак SuperScript II і ThermoScript можуть вступати в реакцію при 50°C або вище, що позбавить від неспецифічних продуктів, що утворюються при нижчих температурах.Для максимальної специфічності суміш РНК/праймер можна перенести безпосередньо з температури денатурації 65°C до температури інкубації зворотної транскрипції та додати до попередньо нагрітої 2× реакційної суміші (гарячий старт синтезу кДНК).Це допомагає запобігти утворенню міжмолекулярних основ при низьких температурах.Кілька температурних переходів, необхідних для RT-PCR, можна спростити за допомогою термоциклера.
3. Зменшує забруднення геномної ДНК:
Потенційна проблема, яка виникає при RT-PCR, полягає в контамінації геномної ДНК у РНК.Використання хорошого методу виділення РНК, такого як реагент Тризол, зменшить кількість геномної ДНК, що забруднює препарат РНК.Щоб уникнути продуктів, отриманих з геномної ДНК, РНК можна обробити ДНКазою I ступеня ампліфікації для видалення забруднюючої ДНК перед зворотною транскрипцією.Розщеплення ДНКазою I припиняли шляхом інкубації зразків у 2,0 мМ EDTA протягом 10 хвилин при 65°C.EDTA може хелатувати іони магнію, запобігаючи залежному від іонів магнію гідролізу РНК при високих температурах.
Для того, щоб відокремити ампліфіковану кДНК від забруднюючих продуктів ампліфікації геномної ДНК, можна сконструювати праймери, які кожен відпалюють для окремих екзонів.Продукти ПЛР, отримані з кДНК, будуть коротшими, ніж продукти, отримані з забрудненої геномної ДНК.Крім того, контрольний експеримент без зворотної транскрипції проводили на кожній матриці РНК, щоб визначити, чи був даний фрагмент похідним від геномної ДНК чи кДНК.Продукт ПЛР, отриманий без зворотної транскрипції, походить від геному.
Час публікації: 16 травня 2023 р



